
Photomicrograph gallery of species of Reduviasporonites Wilson, 1962
a joint initiative with Geoscience Australia
Dr M H Stephenson, British Geological Survey, Keyworth, Nottingham, UK
Prof C B Foster, Geoscience Australia, Canberra ACT, Australia
For this online study, the authors collected and photographed hundreds of specimens of the Permian–Triassic 'fungal' spore Reduviasporonites Wilson 1962 from a wider range of locations than previously documented, including Australia, UK, Saudi Arabia, Russia, Austria and China. The aim of the study is not to detail formal taxonomy, but to use the internet as a forum to display the photomicrographs which show, better than ever before, the full range of variation of the genus.
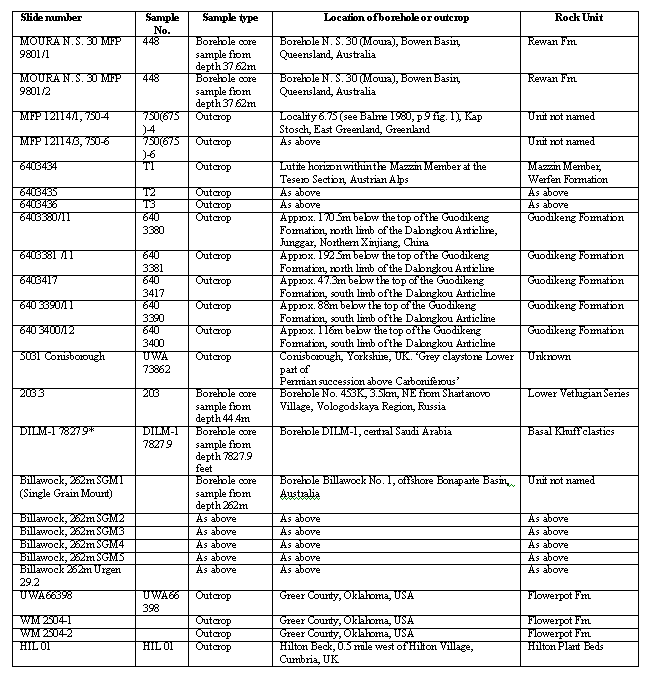
A selection of 242 specimens are presented as a gallery of SEM and light photomicrographs for each of the two species. The gallery, which is searchable, contains brief descriptions of specimens, along with details of specimen locations, slide codes and species determinations. A further table gives details of sample location and type, and the rock unit from which the sample came.


Abstract
Description and illustration of species of the genus Reduviasporonites
(Wilson 1962) Dr M H Stephenson, British Geological Survey, Keyworth, Nottingham, UK.
Prof C B Foster, Geoscience
Australia, Canberra ACT, Australia
A 'spike' of supposed fungal spores occurs at the Permian–Triassic boundary in various locations, in close association with a mass extinction event (Erwin, 1993). This close relationship has lead palynologists (Eshet et al., 1995; Visscher et al.; 1996, Elsik, 1999) to consider that the supposed fungus may have been a saprophytic metaboliser of dead vegetation formed during the extinction event. Elsik recently (1999) suggested that two of the taxa often associated with the 'fungal spike': ChordecystiaFoster 1979 from the Australian Rewan Formation, and Tympanicysta Balme 1980 from the Martinia Shale, Greenland, are junior synonyms of Reduviasporonites Wilson 1962, and affirmed a fungal origin for the latter. It is not clear if topotype material was re-examined to support this conclusion. In contrast Krassilov et al. (1999) suggested that 'spores' assigned by Russian palynologists to Tympanicysta are more closely allied to the zygnemataceaen algae.
For this study, we have examined topotype material of Reduviasporonites chalastus (Foster) Elsik 1999 and Reduviasporonites catenulatus Wilson 1962, as well as well-preserved specimens of these taxa from Greenland, Britain, China, Saudi Arabia, Russia, Australia and Austria, to allow comparative study.
The aims of the project were to:
- describe and illustrate the detailed morphology of Chordecystia Foster 1979, Tympanicysta Balme 1980 and Reduviasporonites Wilson 1962
- determine whether the taxa are synonymous
- determine whether the taxa have fungal or algal origins.
Origin of samples
The specimens described and illustrated in this database are held in the collection of Geoscience Australia. Details given in the database allow each of the specimens figured in transmitted light photomicrographs to be located since slide number and England Finder codes are given for each specimen. For scanning electron photomicrographs the stub number containing specimens is also given.
The slide marked * in the table is not available for examination. Similar material from Saudi Arabia is, however, available for examination from the Palaeontological Collection of the Natural History Museum, London. For details of these slides see Stephenson and Filatoff (2000).
Systematic palynology
Reduviasporonites catenulatus
Reduviasporonites catenulatus Wilson 1962
1967 Reduviasporonites catenulatus Wilson 1962 - Morgan; pl. 1, fig. 1
Description
(Also see Wilson, 1962)
Microfossil, usually forming chains, but also present as pairs of cells or single dispersed cells. Cell outline subcircular or subrectangular; probably originally sub-spherical before compression. Outer cell wall approximately 1µm thick, smooth; may be irregularly thickened at the margins; no inner body or cell material has been observed. Cells in a chain, and particularly single unattached cells, have regular concentric or subpolygonal fold patterns. Method of cell-to-cell attachment not known, though some chains appear to indicate that the convexity of part of a given cell fits into a corresponding hemispherical concavity in an adjacent cell. No terminal rims are present.
Remarks
See Foster et al (in prep.).
Dimensions
See details of specimens.
Range
Permian (for full discussion see Foster et al., in prep.).
Reduviasporonites chalastus
Reduviasporonites chalastus (Foster) Elsik 1999
1979 Chordecystia chalasta Foster: p. 109-110; pl. 41, figs.
3-9; text-fig. 22
Diagnosis
(From Foster, 1979).
Microfossils occurring in chains of two to six individuals or, more commonly as discrete cells. Median members of chain are cylindrical with an elongate rectangular outline in surface view. Terminal members are ellipsoidal to club-shaped. Termini of cells intact; invaginated or convex depending upon position in chain; often slightly thicker and narrower at points of interconnection. Wall one-layered; usually 2µm thick (rarely 1µm). A single fold often extends over the full length of cell; it appears to be a compressional feature. One or two narrow clefts or grooves, each 1–2 µm in width, are apparent on most individuals. The clefts, which may delimit a dehiscence area, extend diagonally between termini of cell or from one end to approximately midway along opposite wall of cell (surface view).
Description
(From Foster, 1979).
Microfossil, usually forming chains but also present as pairs of cells or single dispersed cells. Cell outline subrectangular, rarely oval, irregular or 'Y' shaped, though most cells were probably originally cylindrical in shape before compression. Outer cell wall approximately 0.5µm to 1.5µm thick, smooth; may be slightly thickened in the regions of the terminal rims. The inner body commonly has the same shape as the outer cell and may be adpressed to the inner surface of the main cell so that it is difficult to distinguish optically. The inner body may also occupy a smaller part of the cell cavity (?through shrinkage) and may be twisted. Detachment of the two walls appears first to occur in the middle parts; usually the termini of the inner body are firmly attached to the inner surface of the terminal rim, though detachment at these points may also occur. Wall of inner body 0.5 to 1µm thick where observed, smooth, sometimes hyaline in appearance. Dark 'cell material' of amorphous or granulate appearance occurs in many specimens and is often enveloped by the delicate, shrunken inner body. Cell has a number of characteristic folding patterns. The diagonal folding pattern noted by Foster (1979) is caused by twisting of the cell body about the long axis of the cell. Other folds, in either of the cell walls, commonly delineate rectilinear shapes and these may represent areas of residual attachment of the inner and outer cell walls. Commonly, short lunate folds, orientated perpendicular to the long axis of the cell, occur close to the margins of ovoid cells. These are believed to result from the attachment of the termini of the inner body to the inner surface of the outer body and as such may represent parts of a poorly developed terminal rim.
Remarks
See Foster et al. (in prep.).
Dimensions
See details of specimens.
Range
Permian-Triassic (for full discussion see Foster et al., in
prep.).
Definition of terms
The origin of the forms described in these web pages is uncertain; they have been described as being of fungal origin (Wood and Elsik, 1999; Elsik, 1999; Visscher et al., 1996) and of possible algal origin (Krassilov et al., 1999). In view of this uncertainty a new terminology independent of mycology and phycology is adopted. The new terms suggested are given below (see also Figure 1). For details of the references quoted see the list of references below.
- Cell: the main unit of chains; usually hollow and cylindrical, but may be Y-shaped, ovoid or spherical.
- Terminal rim: the darkened, slightly protuberant annular thickening usually present at the termini or opposite margins of the cell.
- Terminal plane: the usually circular or rectangular area enclosed by the terminal rim, which is perpendicular in orientation to the long axis of the cell.
- Inner body: the thin-walled body, internal to the cell, that normally mimics the overall shape of the cell but which may be of various shapes depending on the extent of its shrinkage.

References
Below are details of all references quoted in the photomicrograph gallery pages.
Eshet, Y, Rampino, M R, and Visscher, H. 1995. Fungal event and palynological record of ecological crisis and recovery across the Permian-Triassic boundary. Geology, Vol. 23, 967-970.
Foster, C B. 1979. Permian Plant Microfossils of the Blair Atholl Coal Measures, Baralaba Coal Measures and Basal Rewan Formation of Queensland. Geological Survey of Queensland Publication, No. 372, 244pp.
Foster, C B, Stephenson, M H, Marshall, C, Logan, G A, and Greenwood, P. in prep. Description, illustration, comparison and biological affinities of species of the genus Reduviasporonites Wilson 1962.
Krassilov, V A, Afonin, S A, and Barinova, S S. 1999Tympanicysta and the terminal Permian events. Permophile, No. 35, 16-17.
Morgan, B E. 1967 Palynology of a portion of the El Reno Group (Permian), southwest Oklahoma. Unpublished PhD thesis, University of Oklahoma.
Stephenson, M H, and Filatoff, J. 2000 Description and correlation of Late Permian palynological assemblages from the Khuff Formation, Saudi Arabia and evidence for the duration of the pre-Khuff hiatus. In Al-Hajri, S. & Owens, B. (eds.) Stratigraphic palynology of the Palaeozoic of Saudi Arabia, Gulf Petrolink, Bahrain. GeoArabia Special Publication No. 1, p. 210.
Visscher, H, Brinkhuis, H, Dilcher, D L, Elsik, W C, Eshet, Y, Looy, C, Rampino, M R, and Traverse, A. 1996. The terminal Paleozoic fungal event: evidence of terrestial ecosystem destabilisation and collapse.Proceedings of the National Academy of Science USA, Vol. 93, 2155-2158
Wilson, L R. 1962 A Permian fungus spore type from the Flowerpot Formation of Oklahoma. Oklahoma Geology Notes, Vol. 22, 91-96.
Wood, G D, and Elsik, W C. 1999. Paleoecologic and stratigraphic importance of the fungus Reduviasporonites stoschianus from the Early-Middle Pennsylvanian of the Copacabana Formation, Peru. Palynology, Vol. 23, 43?53.
Virtual Centre of Economic Micropalaeontology & Palynology (VCEMP)
VCEMP initiates and coordinates the collaborative efforts of micropalaeontologists and palynologists around the world, and, as part of Geoscience Australia's Timescales Programme aims to maintain and develop the Australian national geological time scale with particular emphasis on those parts of the stratigraphic column where economic resources are important.
Present activities
- establishment of a VCEMP/Geoscience Australia Postdoctoral Research Programme
- revision of Campanian biozonation schemes used by industry on the Australian North West Shelf
- global reference studies — linking Australian biostratigraphy to world standard geological stages
- pilot study on Upper Permian conodonts from the Canning Basin
- studies of a proposed global reference section for the continental Permian/Triassic and application to Australian studies
- Sr/Sr dating of Jurassic and Cretaceous shelly fauna from the Australian North West Shelf
- Mesozoic palynology of the Bight Basin.